| Zooplankton Aggregation and Behavior on Ephemeral Fronts in Santa Monica Bay A. Kimo Morris |
updated: 1/07/06
|
|||
|
|
||||
|
||||
 Graduate work of A. Kimo Morris
Graduate work of A. Kimo Morris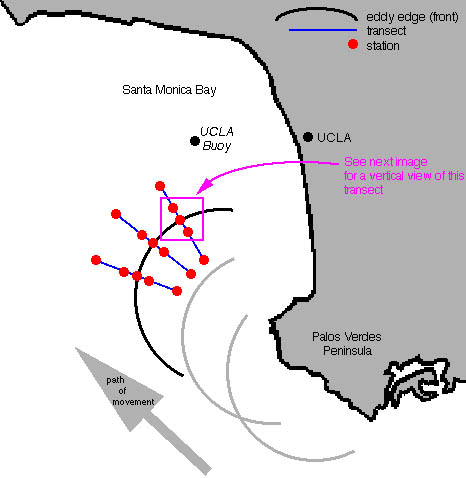 |
|
Click to Enlarge
|
Sampling Goals
![]() I will document differences in the zooplankton community assemblage within the eddy, at the front, and in the water surrounding it. Water contained within the eddy comes from an area outside the geographic region in which it is observed, and it therefore will have physical and chemical attributes similar to the source water but different from that of the surrounding water. As the assemblage of zooplankton is often influenced by physical characteristics of water masses (Franks, 1992), it is reasonable to hypothesize that a difference in abundance, diversity, and community structure of zooplankton groups will also occur between waters within and outside an eddy. The eddies that we are investigating are periodic and ephemeral in nature, and hence, the episodic influx of plankton, particularly larvae, can have a profound impact on the makeup of organism in various habitats within Santa Monica Bay. Documenting the role that recurrent eddies play in larval transport is necessary in order to understand patterns of recruitment of fish and invertebrate populations in SMB.
I will document differences in the zooplankton community assemblage within the eddy, at the front, and in the water surrounding it. Water contained within the eddy comes from an area outside the geographic region in which it is observed, and it therefore will have physical and chemical attributes similar to the source water but different from that of the surrounding water. As the assemblage of zooplankton is often influenced by physical characteristics of water masses (Franks, 1992), it is reasonable to hypothesize that a difference in abundance, diversity, and community structure of zooplankton groups will also occur between waters within and outside an eddy. The eddies that we are investigating are periodic and ephemeral in nature, and hence, the episodic influx of plankton, particularly larvae, can have a profound impact on the makeup of organism in various habitats within Santa Monica Bay. Documenting the role that recurrent eddies play in larval transport is necessary in order to understand patterns of recruitment of fish and invertebrate populations in SMB.
 In particular I will examine the dynamics of zooplankton at the frontal eddy boundary. The ephemeral, topographic fronts seen in the Santa Monica Bay eddy are convergent fronts (DiGiacomo et al., 2002) and thus buoyant plankton aggregates at the surface along the boundary. As a result, planktivores are drawn to the front where food is abundant and accessible. For example, an aggregation of planktivorous red-necked phalaropes was seen by DiGiacomo et al.. (2002), where hundreds of birds were seen aggregated along the frontal boundary of the SMB eddy in a line that was less than 2 m wide. Clearly, transient fronts can greatly impact food web dynamics of Santa Monica Bay.
In particular I will examine the dynamics of zooplankton at the frontal eddy boundary. The ephemeral, topographic fronts seen in the Santa Monica Bay eddy are convergent fronts (DiGiacomo et al., 2002) and thus buoyant plankton aggregates at the surface along the boundary. As a result, planktivores are drawn to the front where food is abundant and accessible. For example, an aggregation of planktivorous red-necked phalaropes was seen by DiGiacomo et al.. (2002), where hundreds of birds were seen aggregated along the frontal boundary of the SMB eddy in a line that was less than 2 m wide. Clearly, transient fronts can greatly impact food web dynamics of Santa Monica Bay. 
Using vertically towed nets, I have begun sampling zooplankton in a grid design across the front. This has allowed a horizontal spatial look at species assemblages and abundance of zooplankton around the front. Concurrent to the plankton sampling, CTD (conductivity, temperature, depth) readings and fluorometer data have been gathered. Latitude and longitude coordinates are recorded. In addition to net-gathered organisms, I have also performed observations on scuba along transects parallel and perpendicular to the front. This will provide information on macrozooplankton that would otherwise not be adequately sampled along the front. I will gather information on species type, abundance and orientation to the front. I have also recorded the presence of sea birds, marine mammals, and anchovy schools in the vicinity of the front.
Lastly, information on concentrations of fish eggs, often seen at fronts (Nakata, 1989; DiGiacomo et al., 2002) will be gathered using a 17’ Boston Whaler boat that will sample fish eggs along the convergent slick simultaneous to the physical and plankton data gathering on the larger research vessel Seaworld UCLA. Neuston nets (0.3 m diameter) will be deployed from booms mounted on the bow of the smaller boat so as to eliminate the disturbance caused by the wake of the boat. One net will be centered in the middle of the slick, thus placing the other net approximately 6 m away from the center of the slick. Three replicate 10-minute tows will be performed with one net on the front. Each replicate will be spaced 0.5 km down the length of the slick. Six additional replicate 10-minute tows will be done, three being performed 100 m inside and three outside the front. Latitude, longitude, speed, start time, stop time, and direction will be recorder. Each net will be fitted with a General Oceanics flowmeter centered in the bottom 1/3 of the nets and these volume measurements will be verified with the latitude and longitude start and stop coordinates. Samples will be fixed in 5% formalin in the field and transferred to 70% ethanol in the laboratory. Volume estimates of each sample will be made, and each sample will be sorted to the lowest reasonable taxonomic level, with special attention paid to larval forms.
Some field efforts have already begun for this project. Thus far, my approach has provided a large, robust spatial database that will be analyzed using parametric and non-parametric correlative tools. Once correlations have been established, I will then utilize spatial autocorrelative geostatistical techniques (as in Coyle and Hunt, 2000) and canonical correlation. Such techniques are appropriate for the data set gathered in this study, since the data have no effective treatment and will be explained across spatial gradients.
 Discussion
Discussion
Data collecting has begun for this chapter, though only rudimentary examination of the data has been done thus far. I hypothesize that the distribution and assemblage of zooplankton including gelatinous macrozooplankton in the vicinity of the edge of the SMB ephemeral eddy is strongly correlated with physical water properties. Depending on the community assemblage at various locations near the front (e.g. at the surface, inside the eddy, outside the eddy, immediately at the front), I hope to address the ecological implications of these transient, although recurrent features of SMB. I will determine (1) how zooplankton assemblages are distributed by the physical oceanographic processes leading to the formation of the SMB eddy, (2) how behavior of plankton accounts for their observed distributions, and (3) how the SMB eddy disperses and aggregates planktonic larvae. In particular, I will assess if the SMB eddy fronts aggregate and transport larvae from offshore back to nearshore parent populations.
- Austin, J. A., and J. A. Barth, 2002, J. Geophy. Res. – Oceans 107(C11): Article 3180.
- Austin, J. A., and S. J. Lentz, 2002, J. Phys. Ocean. 32(7): 2171-2193
- Coyle, K.O., and G.L. Hunt, Jr., 2000, Plank. Bio. Ecol. 47(1): 31-42.
- DiGiacomo, P. M., W. M. Hamner, P. P. Hamner, R. M. A. Caldeira, 2002, J. Mar. Sys. 37: 199-212.
- Franks, P. J. S., 1992, Mar. Ecol. Prog. Ser. 82: 1-12.
- Govoni, J. J., 1993, Bull. Mar. Sci. 53(2): 538-566.
- Graham, W. M., J. G. Field, and D. C. Potts, 1992, Mar. Bio. 114: 561-570.
- Graham, W. M., and J. L. Largier, 1997, Cont. Shelf Res. 17(5): 509-532.
- Kingsford, M. J., and I. M. Suthers, 1996, Estuar. Coast. Shelf Sci. 43: 33-54.
- Le Fevre, J., 1986, Adv. Mar. Bio. 23: 163-299.
- Mackas, D. L., and G. C. Louttit, 1988, Bull. Mar. Sci. 43(3): 810-824.
- Nakata, H., 1989, Rapp. pro.-ver. réun. 191: 153-159.
- Shanks, A. L., 1985, Mar. Ecol. Prog. Ser. 24: 289-295.
- Shanks, A. L., 1988, Fish. Bull. 86(4): 703-714.
E-Mail - Kimo
Other Links
| Academic History | Diving History |
| Kimo's Home Page |
Copyright © 1996-2007, 2008 [A. Kimo Morris]. All rights reserved
Revised: January 7, 2006