|
updated: 03/12/02
|
|
|
My work at Oregon State University was of a theoretical nature. In order to gain experience in life history theory while developing my interest in parasitology, I decided to compare parasites to free-living organisms with regard to life history strategies. In particular, I explored those life-history characteristics of parasites that distinguish them from other organisms, with the objective of inferring evolutionary pathways to the parasitic mode of life. The underlying hypothesis is that life histories can be defined according to both malthusian parameters and species to species interactions, for example, those included within Lotka-Volterra equations. A paradox of parasite life-history is that they combine high reproductive potential with large offspring and long life-expectancy (Calow, 1983). Richer nutrient conditions, compared with predators and grazers, may explain this paradox, however theoretical support is needed to explore its evolutionary origins and redefine the place of parasites within general life-history theory. To address this, I used fundamental concepts in life history theory including trade offs in such parameters as fecudity, generation time, intrinsic rate of growth, and survivorship of juveniles and adults. These variables are usually expressed in the Euler (or Lotka-Euler) equation of general population growth, which can be expressed as:
where lt represents survivorship, mt is fecundity, t denotes generation, and r is the intrinsic rate of population growth. This is a discrete model that accounts for the growth of animal populations that breed in discrete units of time (this model is not intended for organisms that undergo budding or asexual reproduction). Charlesworth (1980) provides the mathematical framework for my approach. If you are truly interested in the derivation, you can email me. Essentially, it leads to a linear equation of the form:
where adult survivorship (lb ) is determined by the negative product of juvenile survivorship (ls ) and fecundity (m) plus the natural rate of population increase (er ). Upon examining this idea, my advisor, Dr. Philippe Rossignol, and I further extrapolated the Euler equation in a way that yielded four extremes in life-history optimization, essentially by introducing an effect on host survival. The principal parameters involved were thus fecundity, adult survival and host survival. Skipping the derivation, the equation takes the form:
The second objective of my project was to state specific evolutionary consequences of this mathematical relationships. To make a long story short, we derived potential pathways of evolution between these trophic extremes, which we named "TROPHIC EVOLUTIONARY PATHWAYS" or TEPs. One important caveat that we made from the beginning was that these trophic strategies required an interaction between two organisms, namely a consumer and a prey item (or host). That meant that we had to throw away all the prior labels that we normally ascribe to animals. For example, fleas are often called "parasites", but our model is only conserned with the ecological effect of a flea feeding on its host. We noticed that the act of fleas feeding on hosts was very similar to aphids feeding on plants. Obviously, both of these examples have some profound differences, but in principle, the ecological effect of both were similar. In both cases, one flea (or aphid) could take meals from multiple hosts, and in every case, the host would not be killed. Remember, were defining feeding strategies, not animals. This meant that we had to come up with new terms for our four extremes. Hence the three letter identifiers in the graph to the right:
We then made the assumption that the evolutionary leap from a free-living existence to an intimate (or parasitic) mode of life would be a one-way ticket, that is parasitic organisms, which exist in a much higher nutrient environment would never increase the quality nor quantity of their reproductive output by evolving a free living existence once again. Thus, once you are parasitic, you will remain in intimate contact with your host. Further, the parasitoid mode of life is highly specialized, and appears to be another one-way ticket to evolution, that is parasitoidism is a terminal strategy. Protelean parasitic wasps are parasitoids that likely evolved from the direction of predation, while certain nematodes (e.g. Family Mermithidae) and other phyla (e.g. Nematomorpha) exhibit an identical ecological effect on their host though likely evolving from the direction of typical parasitism. The convergence on parasitoidism from two different directions is fascinating. The theory is rigorous in that it is clearly refutable from quantitative results; closely related organisms should follow similar evolutionary paths. In addition, widely different species converging on a specific type of relationship could have done so from only two different directions, assessable from quantitative examination. It will also be possible, as a long term objective, to plan experimental studies to further test the hypothesis. This work may contribute important theoretical bases both to parasitism theory, by defining it in a stricter form, and to life strategy evolution by identifying convergencies. E-Mail - Kimo | Wedding '98 | Reception '98 | Hosted by Pukashell.net |
|
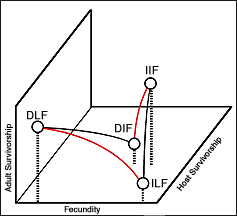 Here all the variables are the same as above with the addition of host survival (lh ). This equations appears to define, in a rigorous way, four trophic strategies - parasitism, parasitoidism, predation and grazing.
Here all the variables are the same as above with the addition of host survival (lh ). This equations appears to define, in a rigorous way, four trophic strategies - parasitism, parasitoidism, predation and grazing. 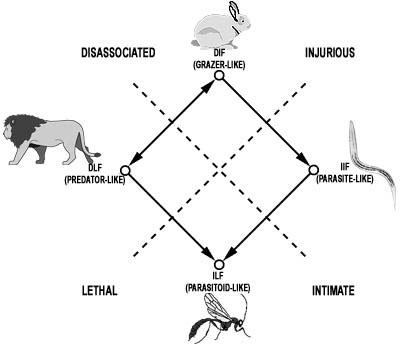 consuming called "parasitoidism")
consuming called "parasitoidism")